AWA: Academic Writing at Auckland
Research Reports may be required in final year or graduate level courses and are usually carried out independently, with the aim to generate new knowledge. A research question is developed, justified and embedded in the existing literature on the topic, and the writer demonstrates strong understanding of research methods and an ability to discuss results and their implications. Research Reports can use topic-specific sections or the standard IMRD (Introduction, Methods, Results, Discussion) sections. They can be presented as dissertations, long essays or research articles (Nesi & Gardner, 2012, p. 136-138).
Title: Relapse in pigeons
|
Copyright: Olivia Salthouse
|
Description: Replicating and extending existing study concerning pigeon behaviour.
Warning: This paper cannot be copied and used in your own assignment; this is plagiarism. Copied sections will be identified by Turnitin and penalties will apply. Please refer to the University's Academic Integrity resource and policies on Academic Integrity and Copyright.
|
Writing features
|
Relapse in pigeons
|
Introduction The resistance of behaviour to change and the extent to which that behaviour will relapse following extinction are two phenomena that have been of interest to behavioural psychologists over many years. The study uses Behavioural Momentum Theory in order to make its predictions about behaviour’s resistance to change, and the extent to which it may relapse after undergoing extinction. Nevin (1995) refers to the basic idea of behavioural momentum by saying that once reinforcement of a discriminated operant has occurred, that discriminated operant will most likely be repeated whenever the stimulus that precedes, it is presented. Similarly, behavioural momentum theory therefore also posits that behaviour will be more resistant to change in a stimulus context that is richer in terms of reinforcement (Nevin, 1995). Response rates and their resistance to change, which form the product of behavioural momentum, are both aspects of behaviour that are independent of one another (Nevin, 1995). This theory has therefore been the subject of numerous studies over the years, and forms the basis for the predictions made in these studies, as well as in the present study. Nevin (1974) carried out a study on resistance to change by conducting multiple experiments to look at the strength of responding in multiple schedules. A multiple VI60s, VI180s schedule was set up and when responding was stable, disrupters, which took the form of pre-session feeding to satiate the birds, extinction and response independent food presentations during the intercomponent intervals, were presented uniformly across the components in the schedule, and the persistence of the responding was measured (Nevin, 1974). The results of this study showed that responding was more persistent in the VI60s component, the richer component, of the schedule, (Nevin, 1974). Such a finding is therefore consistent with the postulates of behavioural momentum theory however, as response rate was also greater in the richer context, these results were confounded and could not prove the independence of response rates and resistance to change in terms of behaviour (Nevin, 1974). Following on from Nevin (1974), Nevin, Tota, Torquato and Shull (1990) undertook a study, which attempted to show that the effects rates of reinforcement and rates of responding had on resistance to change, were independent. In experiment one, a two-component multiple VI60s, VI60s schedule was arranged, with the second of the components receiving 360 supplementary, response independent reinforcers (Nevin, et al., 1990). This experiment showed that the response independent food lowered the response-reinforcer relationship, while increasing the stimulus-reinforcer relationship (Nevin, et al., 1990). A three-component multiple schedule was arranged in the second experiment, which again showed both the response-reinforcer relationship and the stimulus-reinforcer relationship were affected differently by the differing components (Nevin, et al., 1990). Therefore, this study was able to conclude that resistance to change and response rates were both independent in the ways in which they effected resistance to change (Nevin, et al., 1990). A study that furthered research into behavioural momentum theory, by examining relapse in pigeons, was reported by Podlesnik and Shahan (2009). Following baseline sessions and extinction sessions, it was observed that resistance to extinction was greater in the rich component compared to the lean component, which is consistent with the predictions of behavioural momentum theory (Podlesnik & Shahan, 2009). The second finding, of the study showed that relapse, whether it be under reinstatement, renewal or resurgence procedures, was also greater in the rich component (Podlesnik & Shahan, 2009). Again, such a finding is consistent with the statements in Nevin (1995) regarding behavioural momentum theory. This study also corrects the confounding factor of Nevin (1974) by arranging the same response rate on both the rich and lean components. A further study by Podlesnik and Shahan (2010) was conducted, once again looking at behaviour’s resistance to change and the extent of relapse in different components. The results from this study replicated what was found in Podlesnik and Shahan (2009), but also were extended to a number of other species (Podlesnik & Shahan, 2010). The study also furthered Podlesnik and Shahan (2009) by proposing a momentum model of extinction (Podlesnik & Shahan, 2010). Such a model was an extension of the model of extinction that behavioural momentum theory suggested, and was able to explain resistance to extinction and the extent of relapse as effected by differing reinforcement conditions (Podlesnik & Shahan, 2010). This model also fitted previous studies, including Podlesnik and Shahan (2009). Across the numerous studies in this particular subject, it is apparent that the focus of most of studies into relapse, look solely at the internal reinstatement. This means that response independent reinforcers are presented in the components that were previously rich or lean. Little work has been done into external reinstatement where response independent reinforcers are delivered in external conditions, such as the extinction component in the baseline condition (where no reinforcers have previously been presented). Therefore the current study attempts to fill this gap in the literature. The present study was designed to replicate and extend the study of Podlesnik and Shahan (2009), by firstly examining resistance to extinction and then looking at whether relapse of responding occurred following extinction. Provided that relapse did occur, the study also aimed to look at how the history of reinforcement in a particular component affected this relapse. Finally, this study examined whether reinstating reinforcement within one particular component or another component, caused a relapse in responding to occur. This final aim is intended to fill a gap in the present literature by looking at if and how relapse differs when internal reinstatement versus external reinstatement occurs. Based on previous experiments of the same nature, it was therefore hypothesised that resistance to extinction would be greater in the rich component, relative to the lean component. It was also predicted that there would be a greater relapse of responding in the rich component in the internal reinstatement condition. Finally, the study hypothesised that under external reinstatement, relapse would once again be greater in the rich component, in accordance with behavioural momentum theory.
Method Subjects The subjects were 12 racing pigeons, who were maintained at 85% plus 15 grams of their free-feeding body weight, and received post session supplementary food to keep them at this weight. All 12 of the pigeons were experimentally naïve prior to taking part in this experiment and had never pecked a response key before. Each pigeon was housed individually in a separate cage with a 12-hour dark/light cycle. Water and grit were always available in their cages. Apparatus The experiment was undertaken using three pigeon experimental chambers. Each chamber was 295mm in length, 300mm high and 250mm wide. The three plastic response keys all measured 25mm in diameter and were located 190mm above the floor, on the front panel of the experimental chamber, and required a force that exceeded 0.1 N in order to count as a response. There was a 60mm gap between each response key. Only the centre key was used. The hopper, which was located behind the magazine, and raised during reinforcement, was filled with wheat – the reinforcer in this experiment. During food presentations, when the hopper was raised, the magazine aperture was lit and the key lights were extinguished. Food was then made available through an opening in the front panel of the chamber that was 70mm in length and 50mm high, and measured 50mm in depth. The opening was 40mm above the floor of the chamber. The floor of the chamber was constructed of metal rods. At the beginning of each session the house light, which was located in the back wall of the chamber, was lit and then extinguished to mark the end of the session. Also present in each chamber was a camera, connected to a television screen. The television monitors from which the experimental sessions could be observed, were located in the room next door, along with an IBM-compatible PC, running MED-PC software that arranged all experimental events. Procedure Because the pigeons used in this experiment were all experimentally naïve, to begin with they underwent magazine training, which was followed by autoshaping. During the magazine training the pigeons were deprived of food and then trained to eat from the hopper as soon as it was raised. To begin with, the hopper was raised for a full 40-minute session in the chamber. This time was gradually decreased until the hopper was only raised every 15 seconds, for a period of 3 seconds. Once the pigeons were reliably eating food from the hopper as soon as it was raised, autoshaping, outlined first by Brown and Jenkins (1968) began, whereby the pigeons were taught to peck the keys in order to obtain reinforcement. In this procedure, the response key was lit (red, green or white) for 15 seconds, at irregular intervals that averaged 10 seconds. When the light was turned off, food that did not require the pigeon to respond was then presented for 3 seconds. The pigeons started pecking at the lit up key even though a response was not required to receive a reinforcer. When the pigeons started to peck the key the autoshaping became automaintenance, and reinforcement then became contingent on key pecking, which eliminates the light of the key and immediately raises the hopper for 3 seconds. If no peck occurred food was presented after 15 seconds. Our stability criterion for moving the pigeons to the actual experiment required the birds to be pecking the lit up response key 95%, or more, of the time, over five successive sessions. Once the initial training of the pigeons had concluded, the actual experiment began. In order to eliminate order effects, which could render the experiment unreliable and invalid, we counterbalanced the conditions, with pigeons 171 to 176 encountering the internal reinstatement and external reinstatement conditions, in a different order to pigeons 181 to 186. During all of the following conditions, when reinforcement occurred, the hopper was raised for 3 seconds and the magazine light was illuminated for the same 3 second period. In each session, the order of the components was randomly chosen without replacement. Therefore, to begin with each component had a 33.3% chance of being picked and the next component had a 50% chance of being chosen. All 12 pigeons began with condition one, which was a three-component multiple VI 30s, VI 120s, EXT schedule – our baseline condition. The baseline sessions in this condition lasted 40 minutes, or until 40 reinforcers had been delivered, and all birds underwent 30 sessions of this baseline condition. The VI 30s component was the rich context and was signalled by a white key light, the VI 120s component was the lean context and was signalled by a green key light, and the EXT component was the extinction context, signalled by a red key light. Pigeons only received reinforcement when they pecked a white key (rich component) on the VI 30s schedule or a green key (lean component) on the VI 120s schedule of the multiple schedule, but not when they pecked a key that was not lit up or the red key, which signalled extinction. The schedule was presented on the centre key alone. Each component in the schedule was changed after 60 seconds, with a 10 second intercomponent interval (ICI) between each successive component. During the 10 second ICI, the houselight in the chamber was turned off in order to mark a change over in components. The second condition in the experiment, looked at resistance to extinction by instating a multiple EXT, EXT, EXT schedule in which no reinforcement took place in any of the components. Again the white, green and red keys were used, although in this condition they all signalled extinction. The extinction sessions were 40 minutes in length and took place until responding fell to 10% of the baseline response rates. When responding had fallen to 10%, we then moved on to the next condition. In condition three, pigeons 171-176 underwent internal reinstatement, while pigeons 181-186 experienced external reinstatement. In both of these conditions, the sessions lasted 40 minutes in which only three reinforcers were delivered. As with the extinction component, the sessions continued until the bird’s response rates fell below 10% of what they were in baseline. Both these conditions looked at relapse in the pigeons following extinction. In the first session of internal reinstatement the pigeons received three food deliveries that were not contingent on any response, i.e. they were free. While in the first presentation of both the rich and lean components, the food was delivered at 5 seconds, 10 seconds and 15 seconds. The remainder of the sessions in condition three were all arranged as extinction components. For pigeons 181 to 186 who experienced external reinstatement, the same occurred, with three reinforcers that were not contingent on response, being delivered at 5 seconds, 10 seconds and 15 seconds in the first session of this condition only. However the difference was that the food deliveries were made in the extinction component. As with the internal reinstatement condition the remaining sessions in the condition were arranged on a multiple EXT, EXT, EXT schedule. For both groups of birds, condition four was again an extinction condition (the same as in condition two), and again the number of sessions that each pigeon underwent in this condition was dependent on how long it took for their response rates to drop below 10% of baseline response rates. Finally in condition five, we once again compensated for order effects by exposing the two groups of pigeons (birds 171 to 176 and birds 181 to 186) to different reinstatement conditions. This time, birds 171 to 176 experienced external reinstatement in which three food deliveries were made in the first session of the condition at 5, 10 and 15 seconds into the extinction component, and were not contingent on a response. The remaining sessions of this condition were on a multiple EXT, EXT, EXT schedule. Similarly birds 181 to 186 also experienced a reinstatement condition, however for this group of pigeons it was internal reinstatement. In the first session of the condition, the birds in this group were presented with three free food deliveries at 5, 10 and 15 seconds into the rich and lean components. As in the previous reinstatement condition, sessions continued to run until response rates fell below 10% of baseline. All of these experimental sessions were run six days a week.
Results The data that we have collected for this experiment and that is shown below is the number key pecks emitted on the centre key for each of the three components – the rich, lean and extinction contexts that we tested across the baseline condition. In addition, using the proportion of baseline response rates we calculated resistance to extinction and relapse relative to the response rates collected in the baseline condition, these results are also shown. After collecting the raw data from each pigeon, the rate of responding for the baseline condition was calculated by dividing the number of responses by N component. In order to calculate N component, session length was divided by the component duration. For the resistance to extinction and relapse conditions, responding was calculated relative to rate of responding in the baseline condition. This data was then averaged to give the mean rates of responding for the experiment 17 and experiment 18 pigeons. After the analyses were done, the mean data was then plotted to show the average data of the birds. Whilst we set a 10% criterion for the birds in the extinction and reinstatement conditions, whereby session were run until responding fell below 10% of the baseline responding, due to time constraints, this criterion was not used for all of the conditions. The criterion was followed in condition two, but following this we only ran five sessions for the remaining conditions.
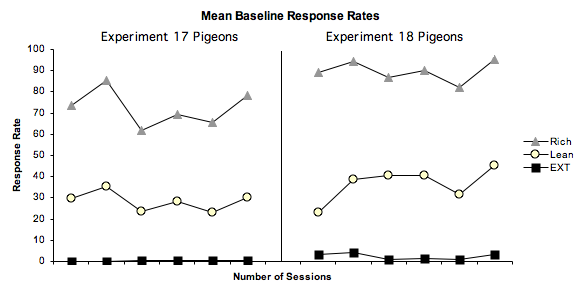
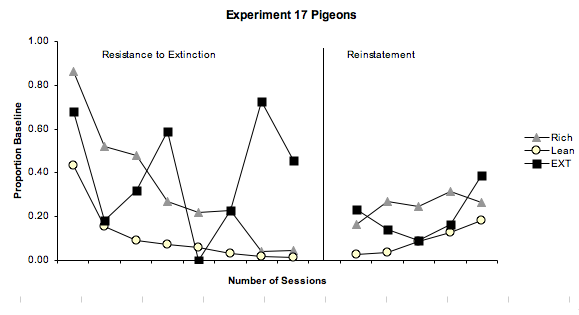
Figure 1. Mean baseline response rates for the experiment 17 and experiment 18 pigeons, in the Rich component (grey triangles), Lean component (yellow circles), and the Extinction component (black squares) of the baseline condition. Figure 1, shows the mean response rate as a function of the session number for the experiment 17 and 18 pigeons during the last six sessions of the baseline condition, to ensure stability of the data. Figure 1 depicts what was expected, with both the experiment 17 and 18 pigeons responding at a greater rate in the rich component than in the lean component. Similarly, in the extinction component we also see what was predicted, with no responding, or at the very least only a very small amount of responding, as with the experiment 17 and 18 birds. Figure 2 shows, for the experiment 17 pigeons, resistance to extinction and relapse as a proportion of baseline responding. The graph showing mean resistance to extinction across these birds shows what was expected for the rich and lean components, with responding, as a proportion of baseline, decreasing over the sessions to below 10%. Also as expected, the rich component shows a greater resistance to change, taking longer than the lean component to fall below the 10% criterion. The extinction component however, does not show a straight line along the x axis as was expected. Instead we see high levels of responding, followed by no responding at all. Such a finding is likely to be related to pigeons 174 and 175 who were not representative of the other birds. In the graph that shows mean relapse in the experiment 17 pigeons, we again see what was expected in the rich and lean components, with the rich component showing a greater amount of relapse than that in the lean component. The extinction component again differs. In this component we see relapse occurring despite no reinforcement being given in this component apart from in the first two sessions, which were the terminal schedule of automaintenance.
Figure 2. The graph on the left shows the mean rate of extinction as a proportion of baseline responding in condition two for experiment 17 pigeons. The graph on the right shows the mean response rates as a function of session number in condition three, the internal reinstatement condition for the experiment 17 pigeons.
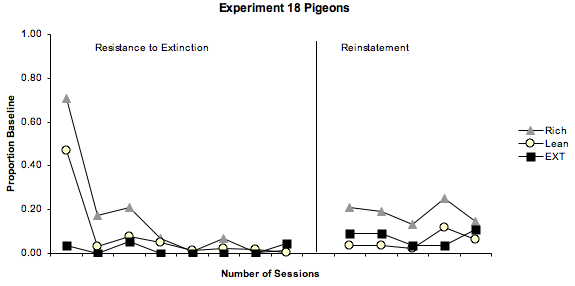
Figure 3. The graph on the left shows the mean rate of extinction as a proportion of baseline responding in condition two for experiment 18 pigeons. The graph on the right shows the mean response rates as a function of session number in condition five, the internal reinstatement condition for the experiment 18 pigeons.
In figure 3, we again see resistance to extinction and relapse as a proportion of baseline responding, this time for the experiment 18 pigeons – minus the data for pigeons 181 and 182, as these birds are still undergoing autoshaping. In the resistance to extinction condition, the rich and lean components again followed a predictable pattern of decrease across the sessions, with behaviour in the richer component taking longer to reach below 10% of baseline responding, relative to the lean component. In the extinction component, the results for the experiment 18 pigeons were a lot more consistent with our hypotheses and with the predictions of behavioural momentum theory, showing from no responding to very few responses at times. In the internal reinstatement condition, relapse followed the predicted patterns in all three of the components, with the richer component showing the greatest amount of relapse, followed by the lean component and then the extinction component in which very little responding occurred at all.
Discussion The results found in this study both replicated the results found by Podlesnick and Shahan (2009) as well as supporting the predictions that were made. The hypothesis that predicted greater resistance to extinction in the rich component compared to the lean component was supported by our results. Similarly, we hypothesised that greater relapse of responding would occur in the rich component in the internal reinstatement condition. Again, our results supported both this hypothesis. Firstly, the finding that resistance to extinction was greater in the rich component, relative to the lean component is consistent with research conducted by Nevin (1974) and Podlesnick and Shahan (2009), who also found that responding was more resistant to extinction in richer components. Furthermore, such findings are supported by behavioural momentum theory, which states that behaviour will also be more resistant to change in a stimulus context that is richer in terms of reinforcement (Nevin, 1995). The reason for this finding is that following the history of reinforcement that had occurred in the previous rich and lean components in the baseline condition, the response rate in the richer component along with a greater amount of reinforcement given, meant that resistance to extinction increased relative to the lean component (Nevin & Grace, 2000). Nevin and Grace (2000) liken this effect to physical momentum in which the product of velocity and mass is momentum. Therefore, a greater rate of reinforcement increases the time it takes for a particular behaviour to be extinguished. Furthermore, the study also found greater relapse under the internal reinstatement condition in the rich component compared to the lean component. Again the history of reinforcement in the rich component was responsible for such a finding, with a greater rate of reinforcement, which when multiplied by response rate gave greater behavioural momentum (Nevin & Grace, 2000). Therefore, this greater momentum meant that when free food deliveries were made in both of the rich and lean components, the pigeons began to emit the behaviour in the same way as they had in the baseline condition, however this was to a greater extent in the richer component. Such a finding is again consistent with behavioural momentum’s predictions of greater resistance to extinction in the richer stimulus context (Nevin, 1995). Likewise these results are consistent with the findings of Podlesnick and Shahan’s (2009) who also found greater relapse in the component that had a richer history of reinforcement. Strangely however, it was also found that relapse occurred in the extinction component for the experiment 17 pigeons, whilst in the internal reinstatement condition, even though reinforcement had never been given in this component. It is possible however, that it was the behaviour that was emitted in the first two sessions of the baseline that relapsed. These first two sessions were the terminal schedule of automaintenance, and therefore the extinction component (as well as the rich and lean components) was run on a VI30s schedule. Whilst a long period of time elapsed between these sessions and the internal reinstatement condition, it is nonetheless possible that it was this behaviour that was seen to be reinstated. One potential limitation of this study was that the total number of free food deliveries in the internal reinstatement condition and the external reinstatement condition, differed. Under internal reinstatement, the pigeons received, a total of six free food deliveries in each session. However, under external reinstatement, a total of three free food deliveries were made. Therefore, the unequal number of food deliveries in these two conditions could have potentially affected our results and thus confounded our experiment and findings. Had the internal reinstatement condition received only three reinforcers in each session, the pecking behaviour may not have resisted change under extinction to the same extent as it did, and it may have been found that internal reinstatement produced differing results than when behaviour underwent external reinforcement. Thus if more relapse was to occur in the internal reinstatement condition relative to the external reinstatement condition, we could not link this finding to the reinstatement condition or the number of free foods delivered overall in the two conditions. Hence such a limitation is a methodological flaw within our experiment. An additional limitation of the current study is the fact that there was a time constraint. As a result of this constraint we were unable to follow the 10% criterion that we had set in place for the extinction conditions as well as the internal and external reinstatement conditions. In condition two, the criterion was followed and the pigeons underwent extinction until their behaviour fell below 10% of what it was in the baseline condition. However, following the second condition, the remainder were only run for five sessions. Had the time constraint not existed and the 10% criterion was able to be followed, it may have produced differing results. As five sessions is relatively short, the behaviour the pigeons had previously learned would not have been fully extinguished and therefore much easier and more likely for the behaviour to relapse. Further, in the internal reinstatement condition, relapse occurred in the extinction component, even though responding whilst in this component was never reinforced. However, the terminal schedule of automaintenance, made up sessions one and two of the baseline condition. During these sessions the key colours were alternated and all were run on a VI30s schedule. Therefore, the extinction component of the experiment initially began as a VI30s schedule, as opposed to in extinction and thus it is possible that when under the reinstatement condition, behaviour from these two sessions relapsed. Such a finding provides evidence for Podlesnick and Shahan’s (2010) suggestion that when behaviour is extinguished it is not ‘unlearned’, but instead just disrupted and therefore can re-emerge under the right conditions. Nonetheless, this methodological flaw is a limitation for this study and would need to be reviewed if the study were to be replicated. Whilst the current study was done using pigeons as its subjects, its findings as well as the predictions of behavioural momentum theory can be extended to the human world. Such findings have implications beyond the lab, particularly in clinical situations when dealing with drug abuse and problem behaviours, which are often link with developmental disabilities. We have concluded that the stimulus context with a richer history of reinforcement produces more relapse when under reinstatement, which can be linked directly to drug addiction relapse as well as problem behaviours. From such results we can conclude that when attempting to extinguish a particular behaviour, relapse of the undesired behaviour will occur to a greater extent in a stimulus context in which there has been a high rate of reinforcement (Mace, et al., 2010). Therefore when training an alternative behaviour to take the place of the behaviour that is undesired, such as self-injurious behaviour, it would be necessary to train this in a differing stimulus context, that is removed from the context in which the problem behaviour exists (Mace et al. 2010). Following the acquisition of the alternative behaviour you could then integrate the stimulus context in which the undesired behaviour occurred (Mace et al., 2010). By doing so, the problem behaviour is receiving much less reinforcement and is therefore much less likely to resist extinction or relapse later on Mace et al., 2010). In the future, looking into the relapse that occurred in the extinction component when under internal reinstatement is a necessary extension of the findings of this current study. Whilst reinforcement was given in the initial two sessions of baseline, there was a fairly long time period between these sessions and the internal reinstatement condition. Therefore, this finding needs to be further looked into to establish for certain that it was in fact the initial baseline sessions that led to this relapse. To do so a novel colour, for example blue, could be used in association with the extinction component, and the amount of relapse that occurred then measured. A further area that these findings could be extended to in the future is drug addiction. While studies have been conducted using behavioural momentum theory’s predictions to successfully eliminate persistent problem behaviours, no research has yet been done to ascertain whether the same principles apply to drug addiction. Therefore a study that looks at training behaviour that is interdependent of drug taking in a differing context from that in which drugs are used may prove to be successful in eliminating this problem. In conclusion, this study examined the theory of behavioural momentum in relation to relapse in differing reinstatement conditions. The results indicated, as we predicted, that resistance to extinction was greater in the richer component of three-component multiple schedule. Similarly the findings showed that relapse occurred to a greater extent in the component with a richer history of reinforcement regardless of whether it was under internal or external reinstatement. These findings are consistent with prior research done by Podlesnick and Shahan (2009), Nevin (1974) and Nevin and Grace (2000) as well as being compatible with the predictions made by behavioural momentum theory. Further research however is necessary to establish our finding of relapse in the extinction component when under internal reinstatement.
References Brown, P.L., & Jenkins H.M. (1968). Auto-shaping of the pigeon’s key-peck. Journal of the Experimental Analysis of Behaviour, 11(1), 1-8. Mace, C.F., McComas, J.J., Mauro, B.C., Progar, P.R., Taylor, B., Ervin, R., & Zangrillo, A.N. (2010). Differential Reinforcement of Alternative Behavior Increases Resistance to Extinction: Clinical Demonstration, Animal Modeling, and Clinical Test of One Solution. Journal of the Experimental Analysis of Behaviour, 93(3), 349-367. Nevin, J.A. (1974). Response strength in multiple schedules. Journal of the Experimental Analysis of Behaviour, 21(3), 389-408. Nevin, J.A. (1995). Contingencies of reinforcement and behavioural momentum: Research and applications. Mexican Journal of Behaviour Analysis, 21, 107-122. Nevin, J.A., & Grace, R.C. (2000). Behavioural momentum and the law of effect. Behavioural and Brain Sciences, 23(1), 73-130. Podlesnik, C.A., & Shahan, T.A. (2009). Behavioural momentum and the relapse of extinguished operant responding. Learning and Behaviour, 37(4), 357-364. Podlesnik, C.A., & Shahan, T.A. (2010). Extinction, relapse, and behavioural momentum. Behavioural Processes, 84, 400-411. |
|