AWA: Academic Writing at Auckland
A Literature Survey provides an overview of the relevant research on a topic. They are often used to build towards a larger research project, such as a Research Report or a dissertation. 3 types of Literature Survey are found in AWA:
An Annotated Bibliography is a list of sources on a topic, usually with the reference details as a heading for each source, and with brief comments on the methods and value of the research in each source. A Research Methods Review reviews research on a topic, but focuses more on the research methods used to investigate the topic (Nesi & Gardner, 2012, p. 148). A Literature Review is like the literature review section of an academic article, dissertation or thesis. The writer surveys existing research on a topic to identify gaps in the knowledge on the topic before explaining their own research. A Literature Review may be presented as an essay or as a chapter.
Title: Chemical morphology of hoverflies
|
Copyright: Stephanie Morton
|
Description: Choose a topic in chemical ecology of insects and write a review.
Warning: This paper cannot be copied and used in your own assignment; this is plagiarism. Copied sections will be identified by Turnitin and penalties will apply. Please refer to the University's Academic Integrity resource and policies on Academic Integrity and Copyright.
Chemical morphology of hoverflies
|
The Chemical Ecology of Hoverflies
Stephanie Morton Corresponding author: Morton, S. (st....@....com)
Key words: Aphid, competition, hoverfly, odour, oviposition, pollination, signalling, syrphid
Abstract Hoverflies (family: syrphidae) are an important prospect for improved aphid control and pollination systems in our agricultural landscapes however, very little is understood about their biology. Hoverflies use a range of environmental cues for foraging and reproduction and chemical cues appear to play an important role in certain aspects of syrphid behaviour such as oviposition, while not being so pronounced in other areas such as nectar and pollen foraging. This review discusses the current literature surrounding the relative importance of volatile odour signalling in hoverflies and how these cues can be utilised to manipulate their behaviour.
Key question: What role does chemical ecology play in hoverfly biology and ecology?
Introduction Insect pollination is a key ecosystem service that the agricultural industry depends heavily on for successful cultivation of many crop varieties (28). There are numerous species belonging to the orders: Hymenoptera, Lepidoptera, Coleoptera, Hemiptera, and Diptera that assist in pollination, with honey bees (Apis mellifera) considered the most important (40). However, with the recent global Varroa mite threat, it has become a concern that relying too much on one species for such a vital service is dangerous (40). Unfortunately, with continual refinement of agricultural crops (increasing yields, pest resistance, and nutritional value), one aspect of crop biology has been left behind, plant pollinator attractiveness (12). As a result there are declining numbers of native pollinators interested in crop species and the dependence on domesticated bees to fill this gap is not only increasing, but beginning to exceed the available commercial supply (1).
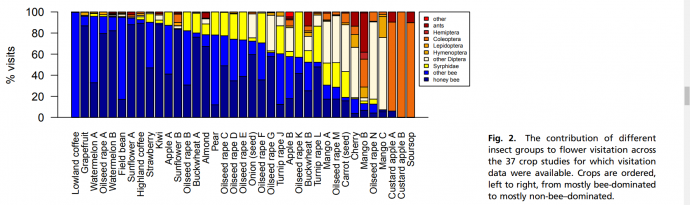
This issue has caused a shift in focus toward a more holistic approach to pollination services, with species diversity and native pollinator recruitment becoming more valued research fields as we try to mitigate the dependence on bees. Non-bee pollinators make up a significant proportion of crop pollinators (see figure 1), and have been shown to assist in increasing fruit set in the presence of bees (17). Syrphidae (hoverflies) in particular have been a vastly underappreciated group (40). Many syrphids not only benefit crops via pollination in their adult form, but feed on crop pests in their larval stages (47). Unlike honey bees (domesticated or wild) aphidophageous hoverflies are not restricted by hive placement or supplementary flower belts, meaning they can uniformly distribute throughout agricultural fields (26). Syrphids are globally abundant and can thrive in agricultural habitats making them an important prospect for enhancing pollination (11; 26; 48).
Figure 1. Ratio of different insect pollinator group’s visitation to agricultural crops averaged across 37 varieties, studied over 5 continents (40).
This review will focus on the recent research that has arisen on the chemical ecology aspects of syrphid biology looking at oviposition, floral, and competition driven chemical signalling and the relative influence these have on hoverfly behaviour.
Hoverfly reproduction Oviposition is a very important function in hoverfly ecology, particularly in the aphidophageous hoverflies where larval fitness and survival depends heavily female choice due to a limited mobility and requirements of aphid prey (5; 8). Females display preferences by altering both oviposition site choice and egg laying rates which can vary significantly across different plant host and prey species (5; 8). What is more remarkable is these biases are also correlated with minor differences in growth rates seen in hoverfly larval instars, indicating female preferences are deliberate and giving their offspring a competitive advantage (5).
A combination of sight and chemical cues aid gravid females in locating and assessing suitable oviposition sites with olfaction working at a distance and sight used in closer (41). Behavioural monitoring of females show that aphid infested plants (with aphids removed) invoke more mobility, searching and oviposition behaviour (19). As little as 500 aphids can be enough for detection of a site by Episyrphus balteatus (the most well studied of the hoverflies), indicating they can be very sensitive to these cues and odours are playing a strong role in eliciting this behaviour (Francis et al. 2005b). Olfactory communication also appears to have a limited importance in young larvae. In another study on E. balteatus 80% of first instars were found to gravitate themselves toward the honeydew from an aphid (Acyrthosiphon pisum) (32). Whereas no preference was observed in second and third instar larvae suggesting as they become older and more mobile they lose the reliance on olfactory senses and use visual cues to detect prey (32).
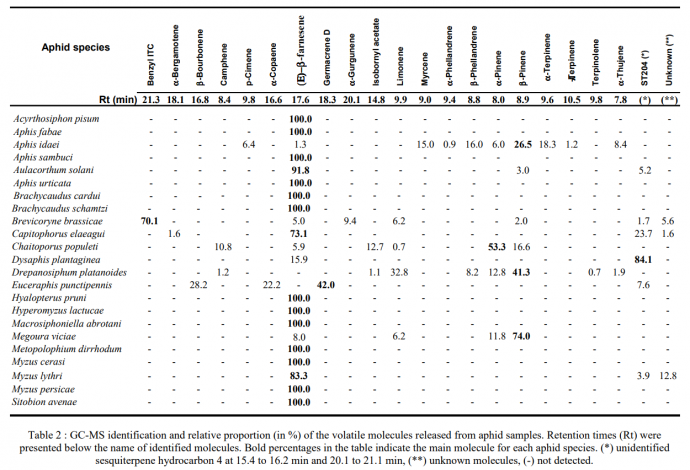
Aphids produce a range of volatile odours that hoverfly may be utilising for detection with E -(β)-farnesene the most prevalent across most species (see Figure 2) (15). α-pinene, β-pinene and limonene are also fairly prevalent in some aphid species such as Megoura viciae and Drepanosiphum platanoides (14). However hoverfly are generally not specific to one compound which fits in line with their generalist predatory nature (14). This research has been taken further with studies displaying artificially induce oviposition using aphid odours, honeydews and sugars as semiochemicals. E - (β)-farnesene was able to increase oviposition significantly while R(+) limonene and (2)-3-hexenol has shown some insignificant trends toward increasing oviposition. Honeydews from aphids can increased egg laying, while fructose and glucose solutions at 30% concentration can produce significant results too, but not sucrose or lower concentrations of sugars (33).
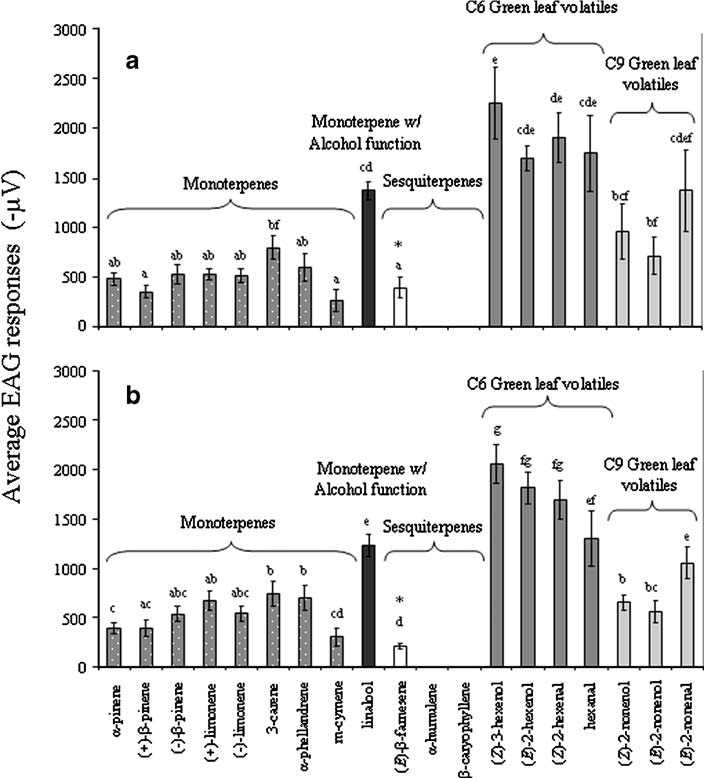
Figure 2. GC-MS relative proportions (%) of volatile odours released from different aphid species. Rt are the retention times (min). Bold percentage indicated main molecule released per aphid species. (*) unidentified sesquiterpene hydrocarbon 4, (**) unknown molecule, (-) not detected (15). Recently there has been new insight from electro antennography (EAG) studies on E. belteatus shedding light onto what compounds aphidophageous hoverflies are capable of recognising from plants and aphids. The most pronounced antennal responses appear to be toward C6 green leaf volatiles (GLV), followed by C9 GLV’s, and then monoterpenes (see figure 3), although it should be noted the relative ratios may be a result of volatility of the substances that have not been corrected for (47). Sesquiterpenes generally do not evoke an EAG response except for E -(β)-farnesene (an aphid alarm pheromone) that females are responsive to, but not males (47). There are a few interesting things to note from these studies. Firstly, prey derived volatiles such as α-pinene, β-pinene and limnonene (15) are recognised by hoverflies, and this supports the notion that they are using aphid prey chemical cues to seek out host plants for their offspring (47). Secondly that many of these compounds are released as herbivore induced plant volatiles (HIPV) and that male hoverflies which are nectar/pollen feeders are responsive to these cues as well. Lastly, males are significantly less responsive to E -(β)-farnesene (the main aphid alarm pheromone) which is unexpected given males are interested in HIPV. Why this so is still unclear in the literature.
Figure 3. Electroantennography (EAG) response activity of females (a) and male (b) Episyrphus balteatus to a range of plant and volatiles (100ug/ml) letters correspond to groupings; bars with no letters in common are significantly different. * indicates a significant difference between male and female response. Five hoverflies were used for each sex and each chemical (47).
Plants have also taken these chemical cues to the next level by developing signals for female hoverflies. Solid-phase micro-extraction (SPME) of potatoes (Solanum tuberosum L.) that have been damaged, undamaged and aphid infested, coupled with gas chromatography-mass spectrometry (GC-MS) analysis found there are 9 volatiles emitted by potato (damaged and undamaged) and these change under aphid herbivory with the addition of 5 supplementary volatiles released; β -sesquiphellandrene, α-zingiberene, β -bisabolene and germacrene D-4-ol, and most notably E -(β)-farnesene (19). β-caryophyllene was also high in potato (damaged and undamaged) but significantly reduced in aphid infested plants. The significance of β-caryophyllene can be speculated from EAG study on ladybug (Coccinella septempunctata) antennae by Al Abassi et al. (2000) that demonstrated that E -(β)-farnesene became less attractive with increasing levels of β-caryophyllene (2). E -(β)-farnesene is thought to be a kairomone released by potato and can also explain the relative attractiveness of potato over other species such as Solanum nigrum that release a mix of monoterpenes, GLV’s and sesquiterpenes but not E -(β)-farnesene (43). Overall the literature is supporting chemical ecology having a strong role in oviposition behaviour.
Foraging and pollination Hoverfly floral visitation tends to support an Optimal-foraging Theory (10). Hoverflies are typically generalist flower feeders (though some can be specialised), focusing on the most abundant and rewarding flowers they can locate (42). Nectar is important for adult survival with visitation rates correlating with nectar quantities (46). Pollen appears less important as a resource except in gravid females indicating the added nutritional content may be required for egg maturation (23; 46). However, floral odour cues are less well understood in the hoverfly and there is little evidence to support its importance.
EAG studies looking at volatile compounds released by plants show a range of plant chemicals that elicit antennal responses in hoverfly’s including monoterpenes and green leaf volatiles (GLF)(45; 47). However, some species are effective at attracting hoverfly pollinators without any floral scent at all. Paphiopedilum barbigerum (Orchidaceae) a deceptive orchid in China is pollinated almost exclusively by female hoverflies, primarily E. balteatus (44) and GC-MS has failed to detect any floral odours in this flower indicating that visual cues of either colour or shape are more important in this instance (44).
Furthermore, in an evolutionary study on Brassica rapa comparing bumble bee (Bombus terrestris) pollination to E. belteatus pollination over multiple plant generations clear selection biases for each species were found (18). Bee pollinated plants developed an almost 2 fold increase in floral odour output. Whereas a significant decrease in three key compounds methyl salicylate, p-anisaldehyde and indole were found in the hoverfly pollinated plants and only one chemical increase, benzyl nitril. Overall hoverfly pollinated plants increased their autonomous selfing leading to inbreeding depression (an expected response to inefficient pollination) which lead to a reduced need for strong floral cues in this species(18). The flowers however, maintained their shape in the hoverfly pollinated population indicating that hoverflies are selecting for cues, but these are visually dominated. These results correlate with a study by Braunschmid et al. (2017) that looked at pollinator species diversity and floral odour variation in an orchid (Cypripedium calceolus) across different geographic ranges in Germany. In Konigsbach, where Syrphidae make a significant proportion of the pollinator species, total scent produced by the orchids is less than half of that (52ng) of Bartholomew (156ng) and Wimbach (134ng) that are dominated by bee pollination (11). In another study Knudsen et al. (2001) also found that palms dominated by syrphid pollination tended to produce floral odours with more compounds in lower quantities than beetle pollinated palms suggesting that they may use odour perfume cues but there is not one particular compound that hoverflies are prioritizing (29).
Overall, these studies suggest that the floral volatile odours in plants are not a vital component to adult hoverfly nectar/pollen foraging behaviour, or certainly not as important as it is to bees and beetles, and that visual cues and plant morphology are still very important for this (20). However, this is not to say chemical cues are completely redundant and to complicate things instances of plant trickery using deceptive floral volatiles have been observed. Head space analysis of another orchid species (Epipactis veratrifolia) found high levels of α-pinene, β-pinene, and β-myrcene in the flowers and trace amounts of α-pinene, β-pinene in the leaf/bud (45). These compounds are also emitted in similar ratios by aphid infested plants and elicit EAG responses in the adult hoverfly E. belteatus (male and females) (45).
To answer the question ‘are visual or odour floral cues more important in hoverfly foraging?’ a study by Primante and Dotterl (2010) compared the two in Cirsium arvense (Asteraceae) flowers and showed that hoverflies were attracted by olfactory cues over visual cues. However after EAG tests they found phenylacetaldehyde, methyl salicylate, dimethyl salicylate, and pyranoid linalool oxide elicited the strongest responses from E. balteatus (39), and although these were prevalent in the floral head space phenylacetaldehyde and methyl salicylate are known to be HIPV’s strongly induced by aphids in other plants (35; 50). Indicating that although chemical cues appear to be important they are heavily associated with aphid infestations rather than pollen/nectar attractiveness qualities. Pyranoid linalool oxide, a common floral volatile (31), does provide one possibility of a floral foraging attractant for hoverfly. This compound and its derivatives are also present in a palm species Prestoea schultzeana that is dominated by syrphid pollination (13; 29) although further research is required to determine if syrphids are exhibiting behavioural responses toward these linalool compounds and how strong these responses may be.
Hetero and conspecific interactions There is very little research into hetero and con specific chemical communication in hoverflies. The only current studies have focused on agriculturally important factors associated with oviposition behaviour such as interactions with other syrphids and ladybugs (Coccinellidae). Both are important factors for biocontrol of aphids with avoidance of predation and cannibalism a desired outcome for optimum biocontrols (3). In studies looking at choice and oviposition in the presence of conspecific eggs, larval tracks (an odour cue) and larvae on plants gravid females will actively avoid plants with conspecifics based on odours alone, and show reduced searching and oviposition as well as greater immobility on these plants when forced into proximity with them (6; 7; 9). Syrphid eggs are also vulnerable to mirid predation (16) and hoverflies have shown aversion to ladybug presence on aphid infested plants. Oviposition is reduced when larval tracks and eggs are detected suggesting chemical cues also play a role in avoiding heterospecific competition and predation as well (7; 9).
So, there does appear to be some chemical signalling utilised by hoverfly although more research is required. Other recent work has found hoverflies will alter their oviposition depending on the parasitism stages of wasps however this is more associated with visual cues. Hoverfly larvae perform best when feeding on un-parasitized aphids but will lay eggs at similar rates on aphids recently parasitized by Aphidius ervi even though there are slight decreases in weight gain, showing they are not as sensitive to this as they are to different aphid or plant species odours (4; 5). Hoverfly will also avoid mummified aphids and exuvia of mummies as their larvae cannot survive on these supporting the idea that visual cues are probably being used (4). Overall, these studies are indicating that hoverflies are recognising some odour cues regarding hetero and conspecific interactions. Unfortunately they have only looked at gravid female hoverfly behaviour and larval growth and survival rates as a response, and this is not representative enough of the whole population to draw conclusions on the relative importance of chemical ecology within these interactions.
Agricultural applications With the current state of knowledge in the field of hoverfly chemical ecology a few potentially exploitable areas of behaviour have become apparent. One is that aphidiphageous hoverflies are using chemical cues for oviposition and the second; these cues are strongly associated with aphid presence. This has prompted the recent development of chemical tools for agricultural use, focusing mainly on HIPV’s as attractive lures mimicking common volatile signals released by aphid infestations. However, different volatile cues are having variable impacts across different species, sex and crop varieties. In a study on three syrphid species (Eupeodes volucris PE, Eupeodes fumipennis, and Scaeva pyrastri) 2-phenylethanol (PE) was found to be very attractive for all three species, but Geraniol and Acetic Acid were only important for E. fumipennis and methyl salicylate for S. pyrastri (L.) (27). A New Zealand study found male and female Melanostoma fasciatum and female Melangyna novaezdondiae were more attracted to rose water lours while male M. novaezdondiae were not (30). Others have found slow release formulations with (E)-β-farnesene and (E)-β-caryophyllene can perform well as attractants for hoverfly, but its effectiveness is dependent on the crop variety it is being used on (22) e.g. 2-phenylethanol is a significant attractant on corn, while eugenol and methyl eugenol are attractant on soybean (21).
Methyl salicylate (MeSA) is the most common HIPV studied across a range of different agricultural crops and insects (24; 35). Using MeSA in field trials on cranberry, soybean, brassica, grapes, and hops it has been found to increase hoverfly trap catch rate in some cases up to 84%, but was not successful in buckwheat indicating there may be a deterrent compound in buckwheat counteracting the MeSA (25; 37; 38; 41). Unfortunately, despite its relative success it appears that MeSA has a very small radius of effect stretching no more than 1.5-2.5m from the lour (36; 41), thus its practical value as an effective lure across large agricultural fields is low.
There are also implications with the measurement techniques of many of these studies. HIPV’s are associated with inducing hoverfly oviposition searching and laying behaviour. This behaviour relies on specific odour cues on the plant, not just a lure, and/or visual cues of aphids when hoverflies arrive in close proximity. This means we cannot infer that hoverflies caught in sticky traps near the lour will also lay their eggs nearby. Two studies that looked at actual syrphid eggs and larvae, and the status of aphids on the plants surrounding a MeSA lour, had conflicting results with one showing greater aphid mortality and the other indicating no increase in hoverfly larvae or eggs (36; 37). This shows that chemical cues can be of use in attracting hoverflies at a distance but there are other important cues used by hoverflies when in close range. This also has spin off implications for the potential of using hoverflies to increase pollination, as HIPV’s are thought to increase oviposition behaviour in females there is little reasoning to expect increased nectar/pollen foraging on the desired crop species especially in smaller patches of crop with alternative floral resources nearby.
Future direction If we are going to utilise these insects in agriculture with chemical manipulation there is still a great deal more we need to understand. A recent genome study by Wang et al. (2017) looked at the antennal transcriptomes of hoverflies and found 154 and 134 transcripts encoding putative chemosensory proteins in E. balteatus and Eupeodes corollae respectively. Most notably two orthologues of volatile sex pheromone detection genes (EbalOBP17 and EcorOBP14) which are known in common fruit fly (Drosophila melanogaster) (49). Another study by (34) found Scaeva pyrastri (L.) has located 59 chemosensory receptor transcripts in adult antennae which is on par with the amount found in fruit fly (Diptera) that exhibits a relatively strong chemical communication system. Thirty seven candidate genes for odorant receptors were found in S. pyrastri (L.)and of those genes, 2 exhibited a male biased expression and 11 exhibited female biased expression meaning there may be yet undiscovered pheromones that could potentially be exploited. These studies indicate that there is still a large amount of information that we do not know about hoverfly chemical ecology. The literature currently has a strong bias toward aphidophageous female hoverflies and from this we have been able to determine that chemical cues are playing a strong role in oviposition behaviour and possibly hetero and conspecific interactions. However, when applying this to practical applications our field measurement methods (sticky traps) need to be revised with stronger focus on monitoring desired behavioural outcomes rather than general presence in an area.
Hoverfly genetics indicate that there may be pheromone cues which are another promising avenue that should be followed with more GC-MS studies on the hoverflies themselves. It is also still speculative as to the role of floral cues in the adult pollen/nectar foraging behaviour and future studies should be focusing on this if we are wishing to use hoverflies for pollination improvement as well as aphid control. To achieve this we need to understand if the volatiles that are attracting hoverflies are being used as floral resource cues as well as aphid signals. The key to this will be in understanding why male hoverflies are attracted to these, are they using them to locate females, food, or both? Secondly, we need more specific testing of key odours that are coming through in the literature as possible floral attractants such at linalool derivatives(29) and benzyl nitril (18). Overall, the recent work on hoverflies has shown us that this is a promising field with a lot of potential but much more research is needed in order to gain a solid understanding of the importance of chemical ecology in hoverfly behaviour.
References
|