AWA: Academic Writing at Auckland
Title: Effects of prey size and spider size on the foraging behaviour in the nocturnal orb web spider Eriophora pustulosa
|
Copyright: Dhobasheni Newman
|
Description: Optimum foraging theory and if/ how it applies to E. pustulosa.
Warning: This paper cannot be copied and used in your own assignment; this is plagiarism. Copied sections will be identified by Turnitin and penalties will apply. Please refer to the University's Academic Integrity resource and policies on Academic Integrity and Copyright.
|
Writing features
|
Effects of prey size and spider size on the foraging behaviour in the nocturnal orb web spider Eriophora pustulosa
|
Abstract There has been little research done on nocturnal orb web spiders. Eriophora pustulosa is a nocturnal orb web spider native to Australia that has been introduced and established in New Zealand. Optimum foraging theory suggests predators should focus on prey items that provide the greatest profit with the least cost to obtain. It has been considered here that this is the case for E. pustulosa. Six sizes of tissue paper balls were made and placed into webs of E. pustulosa and responses toward, away and no response were recorded. There was strong evidence that spider body:ball size ratio and ball size affected the response of E. pustulosa (P=<0.01) and weaker evidence that spider body size affected responded (P=0.03). E.pustulosa went toward smaller balls more often and went away or did not respond more often to larger balls. Further research needs to be done to completely understand the nature of this trend.
Introduction The invention of the spiders web is one of the most innovative and successful designs for prey capture in the natural world (Olive, 1980; Turnbull, 1973). Eriophora pustulosa is a nocturnal orb web spider native to Australia but introduced and established in New Zealand (Authority, 2002). There has been a focus mostly on diurnal orb-web spiders with little work conducted on nocturnal taxa (Ceballos, Henaut, & Legal, 2005) and even less on E. pustulosa. Related species Eriophora edax and Eriophora transmarina are also nocturnal and forage by building a new web each night and dismantling and eating it in the morning before retreating to a location to hide through the day (Ceballos et al., 2005; Herberstein & Elgar, 1994). E. pustulosa was observed to exhibit the same foraging strategy (observations). Optimum foraging theory predicts how a predatory forager would feed most efficiently when given a choice of prey with differing profitability. Predators should therefore focus on prey items that provide the greatest profit with the least cost to obtain (Diaz-Fleischer, 2005). Calorific content of prey has been found to be directly proportional to mass (Suter, 1978) it is logical that predators should focus on larger prey items, but it has been found that larger prey items pose greater cost to the predator based on handling time and the risk of harm in the process (Diaz-Fleischer, 2005; Olive, 1980). Orb web sit and wait foraging is very effective in capturing prey and minimising energetic costs (Olive, 1980; Turnbull, 1973). As this relies heavily on chance of what will come past and get caught it is of benefit to the spider to be able to determine prey items from non-prey and potentially dangerous prey items and the value or calorific content of the prey item to optimise their foraging strategy and prevent being harmed or predated on (Olive, 1980; Suter, 1978; Turnbull, 1973). Studies have shown that orb web spiders are capable of reading information off web vibrations on what is captured in the web and are able to alter their behaviour accordingly (Suter, 1978). The aims of this study are to establish if the size of the spider and prey size have an effect on the foraging behavioural response of Eriophora pustulosa.
Methods Data was collected on 31st of August 2010 between 19:30 and 21:30 in an area surrounding the Miranda shorebird centre located on the firth of Thames to the east of Auckland. It was a dark and cloudy (no moon light visible) night, cold with light breeze following a day of scattered showers. Eriophora pustulosa were located most often on dry fennel, wire fencing and amongst tree branches. Prey ball preparation Six sizes of ball were created out of tissue paper, using 1 – 6 cm2 pieces which were rolled into balls that were equal in size with as much accuracy that could be achieved. Five balls were randomly selected from each size class (1 – 6) then had their diameters measured using high precision callipers to calculate an average diameter for each of the ball size classes. Spider treatment First white light was used to locate and identify spiders as they were often small and there was another spider of similar colour in the same area. After the spider was located and identified as E. pustulosa the white light was turned off and the spider was left for a few minutes (2-3) to ensure any reactions observed were not the result of having white light shone on them. All experimental treatment was conducted under red light. First data was recorded on the substrate on which the spider had built its web, the presence of other food items in the web and whether or not it was currently consuming prey. The size of the spider was estimated by holding a piece of 1x1 mm graph paper as close as possible to 3cm behind the spider to approximate a length using its shadow on the graph paper. Each ball size was tested on 7 spiders and was randomly selected for each spider. Once selected, the size of the ball was recorded then the ball was placed into web in a random location with tweezers and shivered slightly before being released. The spiders response was then recorded (towards, away or no response) in relation to the ball. Data analysis Data was entered into excel and the responses classified at positive (1=towards) neutral (0=no response) or negative (-1=away) in preparation for multiple regression analysis. ANOVA was conducted using the software SAS to produce box plots which was conducted after all preliminary tests were carried out and assumptions met. Multiple regression analysis was conducted using the software R to consider if response was affected by the variables of spider size and ball size after all preliminary tests were carried out and assumptions were met. Results E. pustulosa sits in the centre of its web with its cephalothorax pointing down. When stimulus balls were introduced to the web one of three general responses were observed. The spider either moved away from the stimulus by retreating toward a hub in a corner off the web or rapidly dropping from the centre of the web in a safety line. Or the spider remained at the centre of the web and oriented toward the stimulus or remained did not respond remaining apparently unphased. Lastly the spider oriented and then moved toward the potential prey item. All assumptions were met for statistical testing. The result of the multiple regression analysis revealed 26.2% of the variation to be described by spider size and ball size (R2=0.2622). Though this can considered a weak correlation it was highly statistically significant (P=0.003). table one shows the mean diameters calculated for the ball sizes. Table 1: Mean diameter for ball sizes.
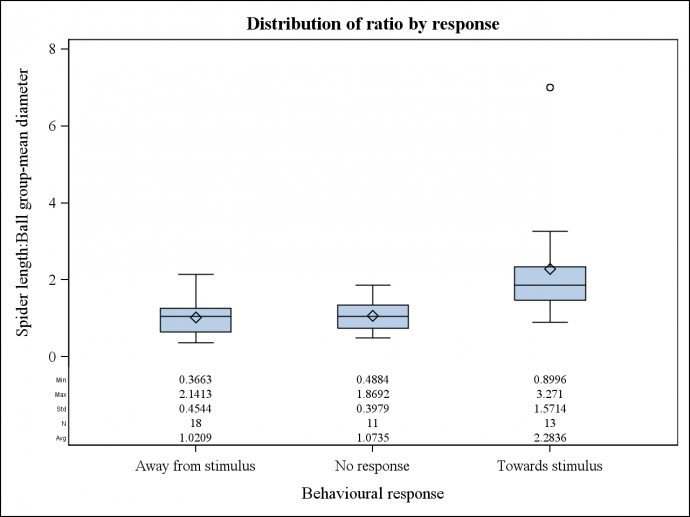
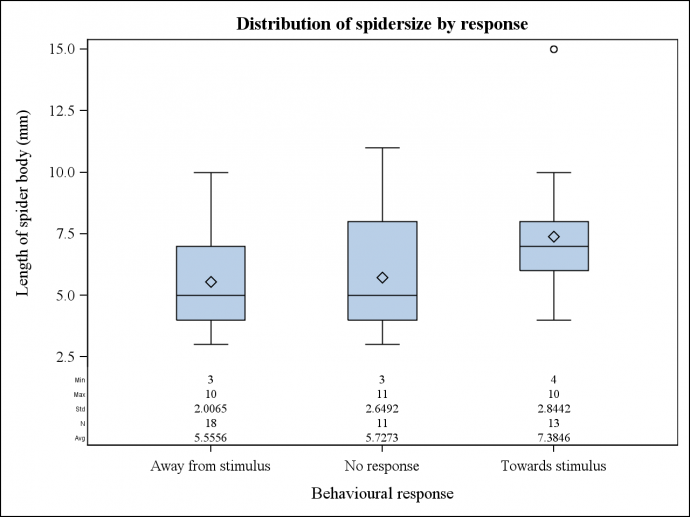
Response as a function of spider:ball size ratio There is strong evidence that the ratio of spider size to ball size has an effect on the likely response of E. pustulosa (F=6.931, P=<0.01). There is evidence here that spiders that were larger than the prey stimulus responded positively more often than those that were smaller. Spiders of similar sizes to prey ball stimulus appear to respond negatively or neutrally on equal occasions (Fig. 1). Figure 1: The effect of body size in relation to prey ball size on the response of E. pusulosa in relation to location of prey. Response as a function of spider size. There is evidence that spider size has an effect on the likely response of E. pustulosa (t=2.257, P=0.030). There is evidence here that spiders that were larger responded positively more often than those that were smaller. Spiders that were smaller (around 5mm) responded negatively or neutrally on equal occasions (Fig. 3). Figure 2: The effect of E. pustulosa body size on response in relation to the location of prey.
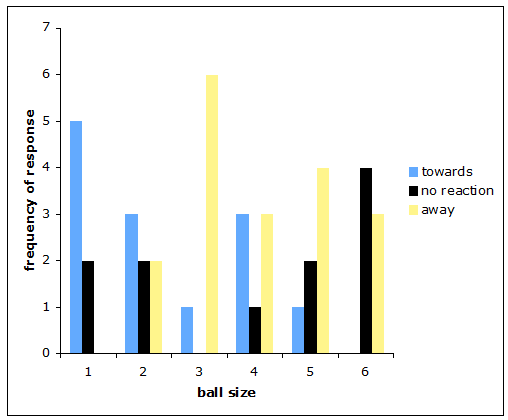
Response as a function of ball size There is strong evidence ball size has an effect on the likely response of E. pustulosa (t=-3.020, P=<0.005). There is evidence here that spiders more often responded neutrally to balls of larger size (>6mm), negatively more often to balls of around 0.5mm and positively more often to balls of smaller size (Fig. 3).
Figure 3: The effect of E. pustulosa body size on response in relation to the location of prey. The frequency with which E. pustulosa responded positively to ball size was higher for smaller balls and negative responses were higher for the larger balls. The highest frequency of positive response was observed for ball size 1 and the highest frequency of negative response was observed for ball size 3. Neutral response remained similar for most ball sizes but frequency was highest for ball size 6 (Fig. 4).
Figure 4: The frequency with which each response was observed for each ball size.
Discussion and conclusions Ball size effects on response There are multiple suggestions in the literature that orb web spiders use web vibrations to establish information about the prey captured in the web (Diaz-Fleischer, 2005; Suter, 1978; Turnbull, 1973). Evidence from this study suggest E. pustulosa is capable of distinguishing between the masses of potential prey. The mass or a prey item is more or less roughly equivalent to its calorific content (Suter, 1978) it can therefore be considered under the optimal foraging theory that single larger prey items would be more profitable than having to catch and process many smaller prey (Diaz-Fleischer, 2005). A study by Diaz-Fleischer (2005) has found that when given a choice Micrathena sagittata will opt for the larger prey item over a smaller one. a similar trend was found by Suter in Cyclosa turbinata. This trend did not appear to be observed for E. pustulosa. Prey items over the ball size of 2 were often avoided or ignored. Under another consideration of optimal foraging theory, larger prey require a greater handling time and therefore a greater foraging energy cost (Diaz-Fleischer, 2005; Suter, 1978; Turnbull, 1973). It therefore may be more prudent for E. pustulosa to ignore these prey items especially as there is always the possibility that they could be inedible or cause harm (Suter, 1978). The negative or retreat response that has been observed can be interpreted as predator avoidance tactics. As orb webs require energy and resource to produce (Turnbull, 1973) it would make sense for spiders to remain with their webs especially as web material is eaten and ‘recycled’ (Ceballos et al., 2005; Herberstein & Elgar, 1994). Therefore behaviours requiring leaving the web (for example drop and retreat behaviour) could be costly to the spider. There have been few studies done considering interactions and predator avoidance behaviour in nocturnal orb web spiders. Many studies on diurnal species consider web decoration and bracing as measures against predators and class being nocturnal as an avoidance tactic (Gillespie & Caraco, 1987; Turnbull, 1973). There are however several potential nocturnal predators who could be feeding on E. pustulosa. Romphaea and Portia are both aggressive mimics whom invade orb webs and mimic prey using an array of complex vibrations to lure in their victims (Jackson, 1995; Whitehouse, 1987). Romphaea uses a sticky web net to apprehend prey (Whitehouse, 1987) whereas Portia is a salticid, pouncing to attack (Jackson, 1995). Both are small in size and are found in either New Zealand or Australia with similar distributions to E. pustulosa (Jackson, 1995; Whitehouse, 1987). Being able to distinguish between a prey item and a mimic potentially has great benefits to the prey spider. E. pustulosa in this study showed greatest frequency of avoidance when treated with ball size three (Fig. 4). Romphaea and Portia are both relatively small predators and there is the potential that E. pustulosa could be avoiding a particular size of prey as a predator avoidance tactic to ensure they are not attacked by predators at any cost. This however is purely theoretical and study needs to be conducted to see if this is the case in New Zealand as only Portia is documented to feed on E. pustulosa in Australia and there is always the possibility that the trend observed is completely by chance due to sampling bias of low sample size. There were strong responses toward ball size one (Fig. 4). The majority of prey observed in webs were small sand flies. The high frequency of responses toward the smaller ball size could be suggesting a preference toward smaller prey sizes that are potentially easier to handle. Argyrodes is a kleptoparasitic spider that uses E. pustulosa as its host (Whitehouse, 1997), stealing prey items from out of E. pustulosa’s web. E. pustulosa has been documented to show aggression toward Agryodes chasing them out of their web and predating on them when possible. Observation of responses to ball size 1 were all rapid and aggressive. Agyrodes is a similar size (2-3mm) (Whitehouse, 1997) to ball size 1 (2.14mm). It is possible that this behaviour is a result of the interaction between the two spiders and E. pustulosa’s attempts to prevent the parasitism. Chasing down Agryodes may be more energetically costly (Diaz-Fleischer, 2005) but the long-term benefits of removing the kleptoparasite from web (by catching and consuming) could potentially out weigh the costs of chasing in that prey will not be stolen in the future. No Agryodes were observed on the webs of E. pustulosa during this study so further research into the nature of this relationship (if it even exists) in the area studied is required. Body size effects on response Spiders of greater body size more often responded toward the prey stimulus. It is possible to use spider size as an index of age and therefore experience, suggesting that more experienced spiders are more capable and likely to attack. Spiders that had a higher body to ball size ratio also responded toward the stimulus on more occasions than those that were smaller. If the spider is bigger than its prey it is likely that the prey will be easier to handle and therefore take less effort to obtain (Diaz-Fleischer, 2005). It can therefore be suggested that this is optimum foraging strategy by E. pustulosa. Though the measurement of body size in this study was not entirely accurate as they were based on the shadow of the spider on graph paper held behind the web, effects observed are real as each measurement estimated was as close to a standard 3cm away from the web as possible. For future replication of this study a more accurate method of measuring the spiders is advised, such as taking photos of each spider which can later be analysed and scaled or calculation of a scaling equation of the size of the shadow to actual size of the spider. There are many factors that could have potentially affected E. pustulosa’s response to the prey stimulus outside the confines of this study. Such as variation in individual experience and condition (Nakata & Ushimaru, 1999; Olive, 1980). It would be interesting in future study to consider if E. pustulosa is able to distinguish between the different objects encountered in its web to establish the state of the arms race between predators and parasites. More observations need to be done on individual spiders by offering a sample of the population a choice between prey sizes and to consider whether experience is a factor in how E. pustulosa responds. There is evidence here that Eriophora pustulosa is capable of determining differences in prey size by web vibrations and adjusts its response accordingly. True to optimal foraging theory E. pustulosa more often went toward smaller prey stimulus that were potentially easier to obtain and avoided those that were larger and could potentially cause harm. Little research has been conducted on this species and further research needs to be done in order to understand the complexity of this interaction.
References Authority, M. B. (2002). MAF Biosecurity pest risk assesment. Spiders accociated with table grapes from United staste of America (State of California), Australia, Mexico and Chile. Wellington, New Zealand: Ministry of Agriculture and Forestryo. Document Number) Ceballos, L., Henaut, Y., & Legal, L. (2005). Foraging strategies of Eriophora edax (Araneae, Araneidae): A nocturnal orb-weaving spider. The Journal of Arachnology, 33, 509-515. Diaz-Fleischer, F. (2005). Predatory behaviour and prey-capture decisionmaking by the web-weaving spider Micrathena sagittata. Canadian Journal of Zoology, 83, 268-273. Gillespie, R. G., & Caraco, T. (1987). Risk-sensitive forrging strategies of two spider populations. Ecology, 68(4), 887-889. Herberstein, M. E., & Elgar, M. A. (1994). Foraging strategies of Eriophora transmarina and Nephila plumipes (Araneae: Araneoidea): Nocturnal and diurnal orb-weaving spiders. Australian Journal of ecology, 19, 451-157. Jackson, R. R. (1995). Cues for web invasion and aggressive mimicry signalling in Portia (Araneae, Salticidae). Journal of Zoology, London, 236, 131-149. Nakata, K., & Ushimaru, A. (1999). Feeding experience affects web relocation and investment in web threads in an orb-web spider, Cyclosa argenteoalba. Animal Behaviour, 57, 1251-1255. Olive, C. W. (1980). Foraging specialization in orb weaving spiders. Ecology, 61(5), 1133-1144. Suter, R. B. (1978). Cyclosa turbinata (Araneae, Araneidae): Prey Discrimination via Web-borne Vibrations. Behavioural Ecology and Sociobiology, 3, 283-296. Turnbull, A. L. (1973). Ecology of the true spiders (Araneomorphae). Annual Review of Entomology, 18, 305-348. Whitehouse, M. E. A. (1987). "Spider Eat Spider": The Predatory Behavior of Rhomphaea sp. from New Zealand. Journal of Arachnologgy, 15(3), 355-362. Whitehouse, M. E. A. (1997). The benefits of stealing from a predator: foraging rates, predation risk, and intraspecific aggression in the kleptoparasitic spider Argyrodes antipodiana. Behavioural ecology, 8(6), 663-667. |
|